
Die an der Lichtreaktion beteiligten Strukturen befinden sich in den Thylakoidmembranen der Chloroplasten. Bei der Lichtreaktion kann man zwei Reaktionen beobachten, die parallel ablaufen. Bei der ersten Reaktion erzeugt ein lichtgetriebener Elektronentransfer NADPH sowie einen Protonengradient. Dabei entsteht als „Abfallprodukt“ Sauerstoff. In der zweiten Reaktion bewirkt der Protonengradient der Erzeugung von ATP (Photophosphorylierung).
Der Aufbau und die Anordnung der Membranstrukturen sind die Voraussetzung für Effizienz dieser natürlichen „Photovoltaikanlage“. Kenntnisse über die strukturellen Zusammhänge wiederum sind die Voraussetzung für unser Verständnis über die Funktion der Lichtreaktion.
![]() Was sind das für Membransstrukturen, die an der Lichtreaktion beteilgt sind und welche Funktion haben?
Was sind das für Membransstrukturen, die an der Lichtreaktion beteilgt sind und welche Funktion haben?
Wie sind sie in der Thylakoidmembran angeordnet und wie lässt sich die Anordnung begründen?
Mehr dazu findet man dann auf der Seite Lichtreaktion und ihre Membranstrukturen – ein vertiefender Überblick.
Der Chloroplast
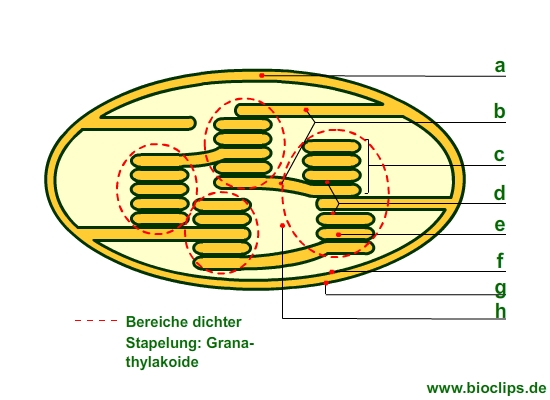
- Chlorplasten haben eine Doppelmembran.
- Zwischen innerer (f) und äußerer (g) Membran befindet sich der Intermembranraum (a).
- Die innere Membran umgibt das Stroma (h): Ort der Dunkelreaktion.
- Thylakoide sind Membranstrukturen (abgeflachte Säcke, Scheiben), die das Stroma ausfüllen und bei den meisten Pflanzen als Grana-Thylakoide (gestapelt) und Stromathylakoide (ungestapelt) vorkommen:
- Granum (c): Scheibenstapel, Bereich dicht gepackter Thylakoide (alle Bereiche dichter Stapelung bezeichnet man als Grana; Grana-Thylakoide)
- Stromathylakoide (b): nicht gestapelte Bereiche, länglich flach
- Stromathylakoide verbinden die Grana – Bereiche.
- Die Thylakoidmembranen trennen den Thylakoidraum (Thylakoidlumen) vom Stromaraum.
- Chloroplasten besitzen drei Membranen: äußere Membran (g), innere Membran (f) und die Thylakoidmembran (d).
- Die Thylakoide entstehen wahrscheinlich durch Einstülpung (Einfaltung) der inneren Membran.
- Die Thylakoidmembranen enthalten die Komponenten für die Lichtreaktion: lichtsammelnde Proteine, die Reaktionssysteme II und I, die Elektronentransportkette sowie die ATP-Synthase.
- Die innere Membran und die Thylakoidmembran sind für die meisten Moleküle und Ionen nicht durchlässig.
- Die äußere Membran ist für kleine Moleküle und Ionen hoch permeabel.
- Das Stroma enthält Enzyme, die im Calvin-Zyklus mittels NADPH und ATP Kohlendioxid in Glukose umwandeln
Die räumliche Anordnung der Photosynthesekomponenten in der Thylakoidmembran
Schulbücher zeigen in der Regel eine lineare Anordnung der an der Lichtreaktion beteiligten Membranstrukturen. Dabei handelt es sich um eine funktionelle Anordnung, die den Reaktionsblauf verdeutlichen soll. In Wirklichkeit sind die Membranstrukturen auf verschiedene Bereiche verteilt. Das hat seine Gründe.
- Die Granathylakoidmembranen enhalten das Photosystem II.
- Die Stromathylakoidmembranen enthalten hauptsächlich das PS I und die ATP-Synthase.
- Der Cytochrom b6f Komplex ist in beiden Thylakoid-Bereichen gleichmäßig zu finden, er kann sich sehr schnell in bzw. zwischen beiden Bereichen bewegen.
- Gleiches gilt für die beweglichen Elektronentransporteure Plastochinon (ist in der Membran frei beweglich) und Plastocyanin (Pc; ist mit Tyhlakoid-Lumen frei beweglich).
- das gemeinsame Thylakoidlumen ermöglicht die Nutzung der vom PSII im Granabereich freigesetzten Protonen durch die ATP-Synthase, die sich weit entfernt in den ungestapelten Bereichen befindet
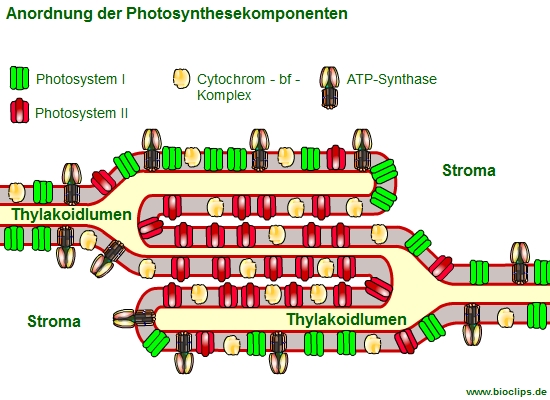
Weshalb befinden sich das PSI und die ATP-Synthase in den ungestapelten Bereichen?
Eigentlich würde man erwarten, dass aufgrund des funktionellen Zusammenhangs PS II und PS I nah beieinanderliegen. Dem ist aber nicht so. Das PS I befindet sich ebenso wie die ATP-Synthase in den Stromathylakoiden.
Der direkte Kontakt zum Stroma erleichtert den Zugang von NADP+ und ADP, auch wäre es für den relativ großen Proteinkomplex der ATP-Synthase in den Granathylakoiden zu eng.
Die dicht gestapelten Granathylakoide sind für das PS II kein Problem, Wasser ist aufgrund seiner Größe ausreichend verfügbar und Plastochinon kann sich aufgrund seiner lipophilen Eigenschaften in der Membran frei bewegen.